�i����j���{�~�ތ�����





��ɊC�y�іk�������m�̌~�ޕߊl�����̐��⌤���ɂ���
2018�N2��2��
��ɊC�y�іk�������m�̌~�ޕߊl�����̐��⌤���ɂ��āiPDF�t�@�C���j
�͂��߂�
���{�̌~�ޕߊl�����ɂ����鉘�������̌����́A��ɊC�ŕߊl�������J�n����1987/88�N�x���s���Ă��܂��B ���ەߌ~�ψ���iIWC�j�ł́A�C��ϓ��Ȃǒn���K�͂̊��ω��ɑ��Đ��E�I�ȊS�����܂������Ƃ��āA1993�N�̔N����ɂ����āu���ω����~�ނɗ^����e���ɂ��Ē������邱�Ɓv���������܂��� (IWC, 1994)�B ���{�́A���̊����ɑΉ����āA���s�̓�ɊC�~�ޕߊl�����iJARPA�j�ɂ����āA�V���Ɂu�~�ނɂ�������ϓ��̉e�����𖾁v���ړI�ɒlj����A1995/96�N�x���i�߂Ă��܂����B
JARPA��2004/05�N�V�[�Y����������18�N�ɂ���Ԓ������I�����܂������A2006�N�ɂ́AIWC�Ȋw���ψ���iIWC/SC�j��JARPA�̍ŏI�]������J�Â��āA����܂ł�JARPA�ōs��ꂽ���������̌������ʂ��R�c����܂����B �����ł́A�N���~���N�N�W���Ɋ܂܂��d�����ދy�їL�@���f�������̒~�ϓ��Ԃƌo�N�ω��ɂ��ĕ�(Fujise et al., 1997)�A�]����̐��ƃp�l���́A���̕����}����ƂƂ��ɁA���Ԋw�I�����y�ё��̓�ɊC�̊����j�^�����O�ɂ���^����\��������Ƃ��ĕ]�����܂����iIWC, 2007�j�B
�܂��A2005/06�N�x����n�܂���������ɊC�~�ޕߊl�����iJARPAII�j�ł́A�u�������������~�ނɗ^����e���̃��j�^�����O�v����ړI�̈�ɉ����܂����B ��ɊC���n���K�͂ł̊������̏I���_�Ƃ��ĂƂ炦�A��Ȋ����������̕��o���ƂȂ�k�����̊C�m�Ƃ̊ԂŌ~�ނւ̉e�����r���邱�Ƃɂ��A�n���K�͂ł̉����ƌ~�ނւ̉e���]�����s�����Ƃ��\�ƂȂ�܂��B �����ŁA���A�̉ۑ�Ƃ��āA�@��ɊC�y�іk�������m�̌~�ނɂ����鉘�������̒~�ςƂ��̐����ߒ��ł̕ω��̉𖾁A�A��ɊC�y�іk�������m�̊C�m���Ԍn�ɂ����鉘�������̋����̉y�сA�B�����������~�ނ֗^���鐶�̉e���̉𖾂��f���Ď��g�݂܂����B 2014�N�ɂ́AIWC/SC����Â��āAJARPA�U�̒��ԕ]������J�Â���܂����B �����ł�2005/06-2012/13�N�̊��Ԃ̌������ʂ���A�N���~���N�N�W���y�уi�K�X�N�W���̐���y�їL�@���f�������̒~�ϗʂ̌o�N�ω��ƌ~�ނւ̌��N�e���]���ɂ��ĕ��܂����B �����2�~��̉��������̑̓��~�ϔZ�x�͑��̒n��Ɣ�r���ċɂ߂Ē�Z�x�ł��邱�Ƃ���A�����̌̌Q�ւ̌��N�e���̉\���͔��ɒႢ���Ƃ���܂����B �]����̐��ƃp�l���́A����ɓ��ӂ��A��ɊC�ɂ���������������̌����́A�~�ނւ̉e���Ƃ����ϓ_����͑��C��Ɋr�ׂėD�揇�ʂ��Ⴂ�Ƃ̌����������܂����B
�k�������m�~�ޕߊl�����iJARPN�j�́A1994�N����1999�N�܂ōs���܂����B ���̒����̎�ړI�́A�k�������m�ɕ��z����~���N�N�W���̌n�Q�\���̉𖾂Ɛۉa���Ԃ̉𖾂ł��B IWC/SC�ł́A�����RMP�i����Ǘ������j��K�p�����Ɏ��g��ł��܂����A�n�Q�\���ɍ��ӂ��邱�Ƃ��ł����A����Ȃ���̎��W���K�v�Ƃ̌������āA�������̎�ړI�Ƃ��Ď��g�ނ��ƂɂȂ�܂����B ���̂��߁A��ʂɂ�DNA�ɂ���`�������p�����܂����A�����⊮����ړI����A�����w�����l�A���w�A�O���`�ԁA�H���A���������A���Ȃǂ̑����삩��̉𖾂����݂��܂����B ���������̒~�ς́A�傽��̓��ւ̎�荞�݂��H���Ɉˑ����Ă���A�����C��������m���݂Ƃ����قړ����C�m���ł��������Ƃ���A�ΏۊC��Ōn�Q���ʂ���Ƃ������_�ŗL�p�ł͂���܂���ł������A���������̒n���I���z�̈Ⴂ�𗘗p���āA�~���N�N�W���̓��̉��������g���̈Ⴂ����~���N�N�W��������Ɋւ���������ȂǁA�n�Q���ʂ̕⊮�I�ȏ��Ɋ�^���܂����iFujise et al., 1997; Nakata et al., 1997�j�B
2000�N����n�܂��������k�������m�~�ޕߊl�����iJARPNII�j�ł́A�u�~�ދy�ъC�m���Ԍn�ɂ���������������̃��j�^�����O�v����ړI�̈�ɉ����܂����i�Ώی~��́A�~���N�N�W���A�j�^���N�W���A�C���V�N�W���y�у}�b�R�E�N�W���j�B �v��̒��ł́A���̖ړI��B�����邽�߂ɁA�@�~�ނɂ����鉘�������i����y�їL�@���f���������j�~�σp�^�[���̉𖾁A�A�H���A���ɂ�鉘�������̐����~�ωߒ��̉y�сA�B���w�I���������ƌ~�ނ̌��N�̊W�̌���3�̉ۑ���f���Ď��g�ނ��ƂɂȂ�܂����B 2016�N�ɂ�IWC/SC��ÂŐ��ƃp�l���ɂ��ŏI���r���[��c���J�Â���A�k�������m�ɂ�����~�ނ̐���~�σ��x���ɂ͕ω����Ȃ����ƁAPCB��L�@���f�n�_�́A���E�I�Ɏg�p���֎~���ꂽ1970�N��ȍ~�A�C�m�����̃��x���������X���������Ă��܂������A2000�N�ȍ~����ԂƂȂ��Ă��邱�Ƃ���܂����B �܂��A����琅���L�@���f�������̒~�σ��x�����Ώی~��̌��N�ɉe����^������̂ł͂Ȃ��ƕ��܂����B ���ƃp�l���́A���N�e�����Ȃ����Ƃɂ��č��ӂ��܂������A�q�Q�N�W���̓��ɒ~�ς��Ă��鐅���L�@���f�������̉�͂ɂ��ẮA�N��Ȃǂŕϓ����邱�Ƃ���A�N���h�{�i�K�Ɋւ�������荞�ĉ�͂��������܂����B ���݂̂����̉�͂�i�߂Ă���Ƃ���ŁA2019�N��IWC/SC�Ɍ��ʂ����\��Ƃ��Ă��܂��B
���̂悤�ɁA��ɊC�y�іk�������m�̌~�ޕߊl�����ł́A�����J�n��������ߊl�Ώی~��̉��������̒������������{���Ă���A����ɂ��ẮA�̓��~�ς̎w�W�ƂȂ�ؓ���̑��A�t���̕��͂�i�߂Ă��܂����i�\�P�j�B �~�̂ɒ~�ς���鐅��ʂ́A������̊������̎w�W�ɂ��Ȃ邱�Ƃ���A�����f�[�^�́A�O�m��̐���̃��j�^�����O�����ɂ��v�����Ă��܂��B �����ł́A����܂ł̒��������̈ꕔ�Ƃ��ē�ɊC�̃N���~���N�N�W���Ɩk�������m�̃~���N�N�W���ɂ��ďЉ�܂��B
�\�P�D��ɊC�̃N���~���N�A�i�K�X�N�W���y�іk�������m�̃~���N�A�C���V�A�j�^���A�}�b�R�E�N�W���̑g�D��������Z�x�ippm ���d�ʓ���j
JARPA�i������ɊC�~�ޕߊl�����F1987/88�`2004/05�j
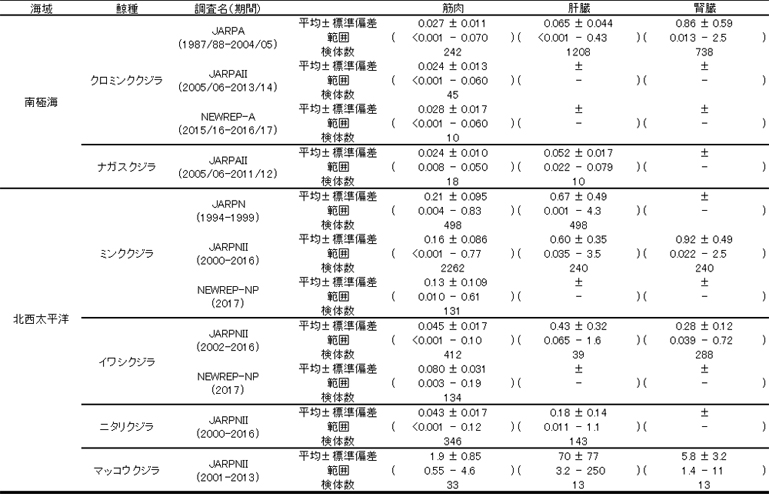
�~�ނ̊̑����̐���Z�x�͒~�ϐ��������A����ƂƂ��ɔZ�x���㏸���邱�Ƃ��m���Ă��܂��iKoeman et al., 1973�j�B �������Ȃ���A���ƕߌ~����i1980/81-1981/82�j�ɓ�ɊC�ŕߊl�����N���~���N�N�W���ł́A20�����Ɍ����X�����݂��܂����i�}�P�j�B ����́A20�ΈȑO�̌̂����炷����Ԃ̉h�{�����ǍD���������Ƃɂ��ƌ����Ă��܂�(Honda et al., 1987)�B
JARPA�Ŏ��{�����~�ނ̊̑����̐���Z�x�͒ʏ�̔N��~�ϐ��������Ă���A�����N�ɂ���Ē~�ϋȐ��̌`���ω����Ă��܂����i�}�P�j�B ����ɁA���i�P�`�T�j�O���[�v�̐���~�σ��x���́A�N�X��������X���������܂����B ���̂悤�Ȓ~�ϋȐ��̌o�N�ω��́A�ۉa���̕ω��f�������ʂł��낤�ƍl�����Ă��܂�(Honda et al., 2006)�B
�}�P�D�N���~���N�N�W���̊̑�������̔N��~�ϋȐ��̕ω�(Honda et al., 2006)
JARPAII�i������ɊC�~�ޕߊl�����F2005/06�`2013/14�j
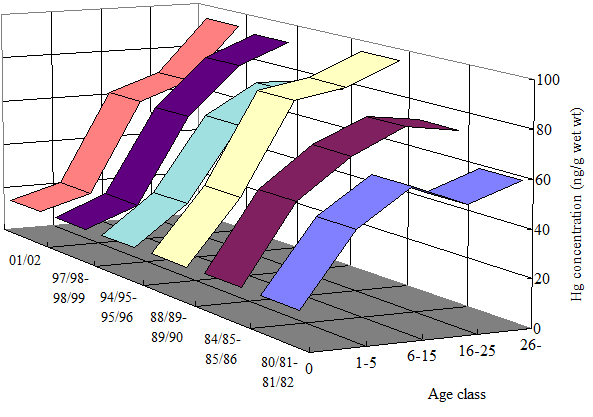
JARPA�̐��ʂƂ��ē�ɊC�̉a���̕ω����A�N���~���N�N�W���̊̑��ɒ~�ς��鐅��̋����ɕω���^���Ă��邱�Ƃ�������܂����B �����ŁAJARPA�y��JARPAII�̊��ԁi1987/88-2010/11�N�j�ɓ�ɊCIV��y��V��i���ەߌ~�ψ���ݒ肵����ɊC�ł̃q�Q�N�W���Ǘ��C��j�ŕߊl�����N���~���N�N�W���̑����̐���Z�x���N���Q���ɂǂ̂悤�Ɍo�N�ω����Ă��邩�����܂����B IV��ł�1988/89�N�ȍ~�A�S�Ă̔N���Q�̐���Z�x�͒ቺ���Ă��܂����i�}�Q��j�B V��ł́A15�Έȉ���26�Έȏ�̔N���Q�̐���Z�x�͗L�ӂȒቺ�X��������܂������A16-25�̐���Z�x�͗L�ӂɏ㏸���Ă��܂����i�}�Q���j�B �����̌��ʂ́A�a�����ɂ߂ėǍD�ł�����1980/81�N����1981/82�N�������ׂāA1987/88�N����̕ߊl�����J�n�ȍ~�A���X�ɃN���~���N�N�W���̉a���������ɓ]���Ă��邱�Ƃ������Ă��܂��B �������Ȃ���AV��̎Ⴂ���n�́i16-25�j�̐��₪�㏸�X���ɂ��邱�Ƃ���A2000�N��ɓ����āAV��̉a���̈��������������邱�Ƃ������Ă��邩������܂���(Yasunaga et al., 2014)�B
�}�Q�D��ɊCIV��y��V��ɂ�����N���~���N�N�W���̔N���Q�i1-15�A16-25�A26�Έȏ�j���̊̑�������Z�x�ippm���d�ʓ���j�̌o�N�ω�
JARPN�i�����k�������m�~�ޕߊl�����F1994�`1999�j
�k�������m�̃~���N�N�W���́A�k�������m��I�z�[�c�N�C����Ȑ�����Ƃ��Ă���O�n�Q�i�I�z�[�c�N�C�]�������m�n�Q�j�Ɠ��{�C�≩�C�A���V�i�C����Ȑ�����Ƃ��Ă���J�n�Q�i���{�C�]���C�]���V�i�C�n�Q�j��2�̌n�Q������܂��B �I�z�[�c�N�C�́A����2�̌n�Q�����݂���C��ł��邱�Ƃ��������Ă��܂��iGoto and Luis, 1997�j�B Fujise et al. (2000) �́A�����n�Q���ʂɉ������������p�ł��Ȃ������݂܂����B �q�Q�N�W���̓��̐���Z�x�͐H���̉e�����h�{�i�K�������ł���قǍ����Ȃ�X�����������ŁA�_��̈��ł���DDT��1970�N��ȍ~���E�I�ɋK������Ă���ɂ�������炸�A���A�W�A�嗤���ł͈ˑR�ꕔ�p�r�Ɍ���g�p����Ă��邱�Ƃ���A���̔Z�x�ɊC�捷�����邱�Ƃ��m���Ă��܂��B JARPN �̐H���������瑾���m���ɐ�������~���N�N�W���͎�ɃT���}�y�уJ�^�N�`�C���V���A�I�z�[�c�N�C�ɐ�������~���N�N�W���̓I�L�A�~����ɐH�ׂĂ��邱�Ƃ��������Ă��܂��iTamura and Fujise, 2002�j�B �~���N�N�W���ɒ~�ς��邱��2�̉��������̊W���r����Ɓi�}�R�j�A�����m���ŕߊl�����̂ɂ͊̑�������Z�x�������̂������A�I�z�[�c�N�C�ŕߊl�����̂ɂ͎��璆DDT�������̂��܂܂�Ă���A�I�z�[�c�N�C�ɗ���O�̐�����̉e�����Ă��邱�Ƃ�������܂����B �����̂��Ƃ���A���{�ߊC�̃~���N�N�W���̌n�Q�����ɁA�����������𗧂\�������邱�Ƃ����炩�ɂȂ�܂����B
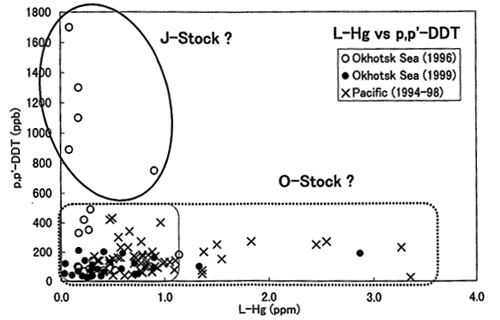
�}�R�D�����m�y�уI�z�[�c�N�C�̃~���N�N�W���̑����̐���Z�x�ippm���d�ʓ���j�y�ю��璆��pp-DDT�Z�x�ippm���d�ʓ���j�̊W��p�����n�Q���ʂ̖͎��} (Fujise et al., 2000)
JARPNII�i�����k�������m�~�ޕߊl�����F2000�`2016�j
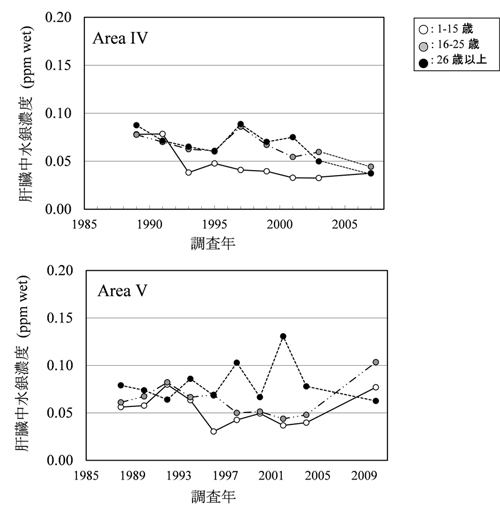
�O�m�ɂ����鐅��Z�x�́A�Y�Ɗv���ȍ~�㏸�������Ă��܂����A���̕ω��͋ɂ߂Ă������Ɛi�s���Ă��邱�Ƃ��m���Ă��܂��B Yasunaga and Fujise (2014) �̕ɂ��ƁAJARPN�y��JARPNII�̊��ԁi1994�`2014�N�j�ł��A�q�Q�N�W���y�т��̉a�����̐���Z�x�Ɍo�N�ω��͂قƂ�NJώ@����܂���ł����B �������Ȃ���CJARPNII�ł��ł������̊C��i9�C��F���o157�x�`170�x�j�ŕߊl���ꂽ�~���N�N�W���̋ؓ�������Z�x�ł́A�o�N�I�ȕω����ώ@����܂����i�}�S�j�B ����́A���̊C��̃~���N�N�W���̈ݓ��e���̒������ʂ���A��v�a�����ł���T���}�y�уJ�^�N�`�C���V�ȊO�ɂ��A����Z�x���ꌅ�����V�}�K�c�I�Ȃǂ����ܕߐH���邱�Ƃ��ώ@����Ă���A���ꂪ�e���������̂ƍl�����܂��B
����́A�C�����̐���Z�x�������C��ɐ������Ă��Ă��A�ۉa�����قȂ�ƁA�q�Q�N�W�����g�̐���~�σ��x���ɂ��e�����邱�Ƃ������Ă��܂��i�}�T�j�B �]���܂��āC�k�������m�̃~���N�N�W���̂悤�ɁC�l�X�ȉa��ێ悷��~��ł́C���̉e�����\�����ӂ���K�v������܂��B
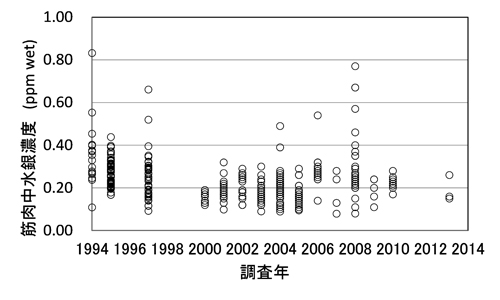
�}�S�D������i9�C��j�̃~���N�N�W���i���n�I�X�j�ؓ����̐���Z�x�ippm���d�ʓ���j�̌o�N�ω� (Yasunaga and Fujise, 2016)
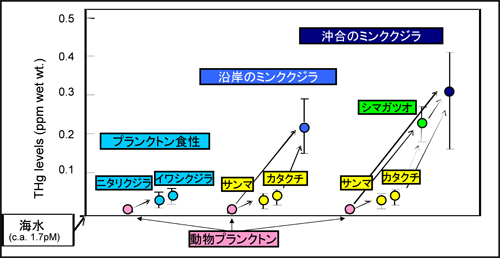
�}�T�D�k�������m�̃q�Q�N�W���ނ̋ؓ�������Z�x�ippm���d�ʓ���j�Ƃ����̉a�����Ƃ̊W��\���͎��} (Yasunaga and Fujise, 2009)
��������
JARPAII��2015/16�N���NEWREP-A�AJARPNII��2017�N���NEWREP-NP�Ƃ��āA�V���Ȓ����v����X�^�[�g���Ă���܂��B ���̐V�����~�މȊw�����ł��A����܂ł̕ߊl�������l�A�~�ނɒ~�ς��鐅�⌤����i�߂Ă��������ƍl���Ă���܂��B
�Q�l����
Fujise, Y., Honda, K., Yamamoto, Y., Kato, H., Zenitani, R. and Tatsukawa, R. 1997. Changes of hepatic mercury accumulations of Southern minke whales in past fifteen years. Paper SC/M97/20 presented to the IWC Intersessional Working Group to Review Data and Results from Special Permit Research on Minke whales in the Antarctic, May 1997 (unpublished). 16pp.
Fujise, Y., Hakamada, T., Aoki, M., Niimi, S., Nakata, H., Honda, K. and Tanabe, S. 2000. An attempt to identify stocks in the western North Pacific minke whale (Balaenoptera acutorostrata.) using the accumulation levels of heavy metals and organochlorines as ecological tracers. Paper SC/F2K/J18 presented to the IWC Scientific Committee Workshop to Review the Japanese Whale Research Programme under Special Permit for North Pacific Minke Whales (JARPN), Tokyo, 7-10 February 2000 (unpublished). 18pp.
Goto, M. and and Pastene, L.A. 1997. Population structure of the western North Pacific minke whale based on an RFLP analysis of the mtDNA control region. Rep. Int. Whal. Commn 47: 531-537.
Honda, K., Yamamoto, Y., Kato, H. and Tatsukawa, R.1987. Heavy metal accumulations and their recent changes in southern minke whales Balaenoptera acutorostrata. Arch. Environ. Contam. Toxicol 16: 209-216.
Honda, K., Aoki, M. and Fujise, Y. 2006. Ecochemical approach using mercury accumulation of Antarctic minke whale, Balaenoptera bonaerensis, as tracer of historical change of Antarctic marine ecosystem during 1980-1999. Bull. Environ. Contam. Toxicol 76: 140-147.
International Whaling Commission. 1994. Chairman's Report of the Forty-Fifth Annual Meeting, Appendix 12. Resolution on research on the environment and whale stock. Rep. int. Whal. Commn. 44: 35.
International Whaling Commission. 2007. Report of the Intersessional Workshop to Review Data and Results from Special Permit Research on Minke Whales in the Antarctic, Tokyo 4-8 December 2006, Paper SC/59/Rep1 presented to the IWC Scientific Committee, May 2007, Alaska (unpublished).
Koeman, J.H., Peeters, W.H.M., Koudstaal-Hol, C.H.M., Tjioe, P.S. and de Goeij, J.J.M. 1973. Mercury-selenium correlations in marine mammals. Nature 245: 385-386.
Tamura, T. and Fujise, Y. 2002. Geographical and seasonal changes of the prey species of minke whale in the northwestern Pacific. ICES Journal of Marine Science 59: 516-528.
Yasunaga, G. and Fujise, Y. 2009. Additional analyses of temporal trends and factors affecting mercury levels in common minke, Bryde's and sei whales in the western North Pacific, Paper SC/61/JR3 presented to the 61st IWC Scientific Committee, May 2009 (unpublished). 21pp.
Yasunaga, G., Bando, T. and Fujise, Y. 2014. Pattern of mercury accumulation in the Antarctic minke whale and its prey based on JARPAII data. Paper SC/F14/J23 presented to the JARPA II special permit expert panel review workshop, Tokyo, February 2014 (unpublished). 7pp.
Yasunaga, G. and Fujise, Y. 2016. Temporal trend of Total Hg levels in three baleen whale species based on JARPNII data for the period 1994-2014. Paper SC/F16/JR30 presented to the JARPNII Review Workshop, Tokyo, February 2016 (unpublished). 14pp.